Muscle
Structure and Function
Muscle Fiber
Types
Muscle growth
Lactic Acid
Muscle tissue can be one of several types in the body: smooth, cardiac, or skeletal. For our purposes we are concerned with skeletal muscle, which makes up the bulk of the body's muscle and is the tissue we use for physical activity.
The muscle that you can see is composed of subunits called fascicles. Fascicles are bundles of individual muscle fibers. Each fiber is one elongated cell that may extend for the length of the muscle. Each muscle fiber cell has several nucleii (unlike most cells, which have only one), and is segmented into distinct sectional bands. Within each muscle cell are numerous myofibrils, which also extend for the length of the muscle cell. Sarcomeres are the basic contractile subunit of myofibrils.


Actin and myosin are the two principal muscle proteins, and they are found in myofibrils. They are arranged in a ring-like structure, usually with six (thin) actin strands surrounding a (thicker) myosin fibril. Again, they run parallel and lengthwise. The myosin fibril has numerous small protrusions called crossbridges. The actin strand is actually intertwined with an even thinner, ribbon-like protein called tropomyosin, and a smaller molecule, called troponin, associates with tropomyosin in this structure.
When a nerve impulse signals the muscle to 'do something' the activity causes channels in the sarcoplasmic reticulae to open their gates and release calcium into the cytoplasm. In skeletal muscle cells the sarcoplasmic reticulae (SR) is an extensive intracellular network which serves as the storage area for calcium. Usually, the inside of the cell cytoplasm has a very low calcium concentration. When it rises, some of the calcium diffuses over to the muscle protein fibers and causes a conformational change.
Ca++ ions will bind to troponin and cause it to rotate slightly. This is enough so that the tropomyosin moves, and actin now is exposed to the myosin below. The myosin crossbridge then jumps up and binds to the exposed actin. The myosin crossbridge drags along the actin fiber like a ratchet, completing a "power stroke".
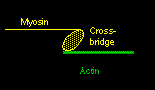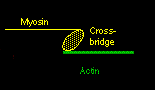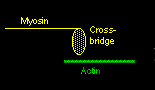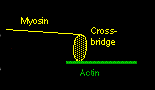
When all the crossbridges in a sarcomere do this at the same time, the sarcomere contracts. After the nerve impulse ends, the SR has mechanisms to reabsorb the free calcium and put it back into storage. As calcium disassociates from the troponin, ATP binds to the crossbridge to 'disconnect' the bridge from the actin. The actin fibers change back to their previous positions and the sarcomere relaxes.
A muscle cell does not necessarily go back to complete relaxation right away. It can remain contracted through a series of stimulations. This process, called summation, increases the total force of muscular contraction. When the stimulus is great enough, many sarcomeres in many fibers are "recruited" and the muscle as a whole contracts. This is why we can lift or push varying amounts of resistance . . . more or less cells are recruited, and to a greater or lesser extent. Muscle failure occurs at the point where the maximum number of fibers are being stressed to their limits.
If you realize that each crossbridge requires ATP, and each myosin strand has dozens of crossbridges, and each muscle fiber has hundreds of myosin strands, you can see that muscular exertion requires a lot of energy. Glycolysis provides some energy (ATP), but the real engine is the Krebs cycle, which requires oxygen. As available oxygen diminshes during muscular work, incoming pyruvate *ferments* into lactate (aka lactic acid) instead of getting converted into acetyl coenzyme A. Lactate travels out of the cell and through the blood to the liver, where it may be reconverted to pyruvate and glucose and return to the cell, in a process called the Cori cycle. Or the blood lactate can enter a different muscle cell and get reconverted to pyruvate (and eventually acetyl coA) if there is sufficient oxygen. However, intracellular levels of lactate can rise more quickly than it can be carried off and this results in a painful local burning sensation that requires a temporary cessation of muscular work. This is the well-known "burn" of weight training.
Given the huge energy needs of muscle, and its importance to the organism's survival, the body also has a backup system. ATP in muscle exists in equilibrium with creatine. After ATP gives up a phospate group and becomes ADP, creatine phosphate will give it a phosphate group and regenerate ATP. This can allow muscular work to continue even while more ATP is being created in the Krebs cycle.
Creatine is a short-term yet important reserve . If it is available to recharge ATP, the cell will not need to resort to glycolysis to make more in the immediate term. So what? Well, remember that the muscle is working hard, and the Krebs cycle is running at full blast. Oxygen is in short supply, so making ATP from glycolysis at this point would be anaerobic, meaning fermentation and lactate buildup. Creatine cuts the muscle some slack, and it can keep working longer before exhausting the ATP and hitting "the burn".
Thanks to Tom Burkholder and A. Murray for clarifications.
Muscle Fiber
Types
With regards to the muscle cells there are two types of fibers.
Type I- also previously known as slow twitch. These fibers are only aerobic and contain an abundance of mitochondria (where the Krebs cycle occurs). The major source of energy for Type I is fat through the Krebs cycle. These are the muscle fibers we continuously use throughout the day for walking etc. Low intensity work. These are also the muscle fibers which are used during activities which are moderate in intensity, up to 75% max heart rate. Most would call this cardio work.
Even within these muscle cells the first stage of glucose breakdown is anaerobic with the formation of two ATP + pyruvic acid. The pyruvic acid then enters the mitochondria to be further broken down in the Krebs cycle.
Type II has three subgroups:
a- Fast Oxidative Glycolytic- These fibers are also aerobic and contain some mitochondria. With proper training (Anaerobic Threshold training) they will increase the number of mitochondria. The major source of energy is glucose. Most of the literature that I have read talks about glucose as being the only source of energy for these muscle fibers. I suspect that under certain circumstances they may eventually adapt and burn fat as well but I can find nothing that backs this concept. These fibers come into play when the intensity of our work is between 75% and 85% max heart rate (AT type training).
b- Fast Glycolytic- These fibers are anaerobic. They contain no mitochondria and do not have the enzymes for the Krebs cycle. Their only source of energy is glucose. They produce an abundance of lactic acid. They come into play when intensity of work is in the anaerobic range. High intensity activities like sprints, high intensity weight lifting or powerlifting.
c- Indeterminate- may become type a or b depending on the type of training.
Those athletes who are genetically adapted to become very muscular - weightlifter types and body builders - are likely to have 40% Type I and 60% Type II whereas those destined to become great endurance athletes probably have 60-80% Type I and 20-40% type II muscle fibers.
During aerobic exercise when the primary muscle fibers being utilized are type I, FFA are used even in the presence of adequate glucose. Generally during activities of intensity between 60-75% max heart rate the primary muscle used are type I. As intensity increases (to 75-85%), Type IIa come into play more and more and they utilize glucose as their source of energy-these fibers produce most of the lactic acid. Aerobic exercise intensity between 70-80% max heart rate utilizes both type I and type IIa fibers. Aerobic exercise intensity between 80-85% uses almost exclusively type IIa fibers. Exercise intensity above 85% utilizes only type IIb fibers- anaerobic. When you run out of glucose you hit the wall and the only sustainable exercise is in the 60-70% range-type I fibers.
Type I- aerobic- burns mostly fat- lots of mitochondria- not lactic acid- need oxygen.
Type IIa- aerobic-burns glucose-mitochondria present but not as many as type I
Type IIb- anaerobic- burns glucose- no mitochondria- produces lots of lactic acid which can enter mitochondria of type I or IIa - heart muscle can utilize all the lactic acid it can get. Any lactic acid left over enters the liver and is reconstitued into glucose.
Type IIc-undifferentiated fibers -can become type IIa or IIb.
Note: it would take a lot of exercise to deplete the glycogen stores in muscle. Since even high intensity aerobic exercise uses in the vicinity of 15-20 cal/minute it would take at least 60 minutes of high intensity work to depelete glycogen stores.
Endurance athletes learn to pace themselves so as to utilize both type I(fat burning) as well as type II (glucose burning) fibers and leave enough for the final sprint (type IIb).
Glucose drinks help spare the glycogen and are needed only in events lasting more than 90-120 minutes.
Note: weight lifters use primarily type IIa and type IIb muscle fibers. Perhaps that is why they have so much trouble getting rid of excess fat despite a lot of work. If weight lifters would incorporate more aerobic workout they would have less to worry about when it came to body fat. Not too many years ago most endurance athletes would cringe at the suggestion of using weight lifting to enhance their performance. Now it is used religiously by elite endurance athletes (especially in the off season) without sacrificing their performance.
I hope that I have been of help.
Ralph Giarnella MD
From: mikeprevost@net-star.net (Prevost, Mike)
Subject: Re: Muscle Fiber Type Conversion
Several studies have shown the conversion of type II fibers to type I.
Under conditions of chronic electrical stimulation the Extensor Digitorum Longus in rats (which is primarily fast twitch) becomes almost entirely slow twitch, indicating a considerable conversion of type II to type I. Also, hyperthyroidism in rats can convert the almost entirely slow twitch soleus muscle in rats to primarily fast twitch. If we immobilize a limb we can also observe a conversion of fast twitch fibers to slow twitch fibers. In fact, there are a host of different treatments (drugs, metabolic perturbations, hormones, activity patterns) that can enduce a large scale muscle fiber type conversion. The fact that we can shift a muscle from 98% type I to better than 50% type II indicates that some fibers are shifting from type I to type II.
The conversion probably proceeds from type IIb to type IIa to type IIx then type I. In fact, evidence now indicates that the fast twitch contractile proteins (type IIa and probably also IIb) are constitutively expressed (expressed by default in the organism) and that it takes some kind of stimulus to express the type I contractile proteins. This is supported by the numerous studies on muscle adaptions to disuse. If muscles are not allowed to contract (eg, by severing the nerve or physically immobilizing the limb) the muscles begin to express the fast contractile proteins. The longer the immobilization, the more fast twitch the muscles become.
Dr. Giarnella is correct to the extent that it has been difficult to demonstrate a conversion of type I fibers to type II due to exercise training. The current theory proposes that type IIb fibers are converted to type IIa during strength training and type IIb are converted to IIa and type IIa to type I during endurance training programs. Years ago it was assumed that muscle fiber type was fairly stable and genetically determined. This has been disproven in recent years by literally hundreds of studies indicating that muscles posses an incredible degree of plasticity.
Mike Prevost, Ph.D.
Muscle Growth
Muscle growth is a specialized form of protein synthesis. As we saw above, a steroid hormone (testosterone) enters the muscle cell by diffusing directly across the cell membrane, combines with a receptor in the cell and then stimulates gene transcription and protein formation via the DNA -> mRNA -> tRNA -> protein pathway. Specific receptors and genes are involved.
Muscle cells, as mentioned before, are long cells called myofibrils. They differ from most other cells in that when muscles grow, the individual cells simply become thicker and longer instead of dividing into entirely new cells. Muscle cells also differ from most other body cells in that muscle cells are multinucleated. A myofibril may increase in size up to 28 times its initial size.
The interesting questions come in as we start looking at exactly how and when this process occurs. Human growth hormone (hGH) and insulin-like growth factors (IGFs) seem to play an important, though somewhat unclear, role.
hGH is released from the anterior pituitary and travels through the blood. It acts on the liver to release IGFs. Both IGFs and hGH are peptide hormones; IGFs are structurally very similar to a large section of the insulin molecule - hence their name.
What precisely happens at the muscle cell is not known, but we can make some fairly well-informed speculation. Since IGFs are similar to insulin, it makes sense to think that they would also have a similar function. So IGFs probably work to increase uptake of amino acids and glucose into muscle cells. It is not clear whether muscle cells have receptors for hGH, but if they do, then it could be that hGH increases nuclear division in muscle without triggering cellular division (mitosis).
We have seen how DNA and RNA are critical to protein synthesis, so it is clear that having more nucleii within muscle would be very beneficial for more rapid protein synthesis (muscle growth). It turns out that each nucleus has a sort of effective "range". When the muscle grows, it can only grow as far as the nucleii will "reach". So the number of nucleii control the ultimate size of the muscle fiber. One of the major functions of hGH is to stimulate cell division. Now, if there are hGH receptors in muscle, but muscle cells lack the ability to divide, and hGH has an anabolic effect on muscle, it stands to reason that hGH is increasing the nuclear division process (and thus the total number of available nucleii in the muscle), but the cytoplasmic separation process never kicks in. Perhaps the mechanism for it that is found in most cells has been lost over time in muscle as an evolutionary adaption. (There is no doubt that muscles are very important to survival!)
It seems then that hGH and IGFs might have complementary functions in stimulating muscle growth. hGH could be instructing the muscle cells to "build more factories" for muscle while IGFs could be stimulating the cells to take in more "building blocks" for protein synthesis. Both hGH and IGFs may affect other important components in the process as well - such as increasing the production of hormone receptors or tRNA or activating enzymes that accelerate transcription.
Multinucleation might explain the longstanding anecdotal phenomenon most bodybuilders call "muscle memory". Muscle memory is recognized when someone who has had a substantial muscular mass and then lost it due to injury or layoffs from training, returns to training and regains the majority of the mass in a much shorter time than was initially required to develop it. What could be happening is this: the specific muscle proteins in the muscle were cannibalized by the body for energy production during non-use. However, the muscle retains the higher than average number of nucleii that the previous exercise stress caused the body to create. When presented with exercise and proper nutrients, new protein synthesis can occur at an accelerated rate.
Copyright notice: The entire contents of this document are Copyright 1997 Paul L. Moses. Redistribution for personal, noncommercial use is permissible, so long as the document is distributed in its entirety, including this notice. Any other use requires an express written request and consent from the listowner. All rights reserved.
Lactic Acid: Effects on Muscle Function
"Muscle Memory": A Theoretical Concept Based on Athletes' Reports
ECA (Ephedrine, caffeine and aspirin) stack
